OISTの博士課程に所属して3年目になったので、これまでの流れをまとめてみます。
目次
OISTに入学する前の準備などはこちらの記事に書いています。
-
主な登場人物
- メインの指導教員 (Thesis Supervisor): ローテーション終了後から4年間所属する研究室のPIの先生を指します。
- メンター (Academic mentor): 在学中に何でもアドバイスをしてもらえる先生です。
- ローテーションの指導教員: ローテーション期間中にお世話になる先生です。
- Co-supervisor またはThird Committee Member: あなたの研究についてアドバイスをくれる3人目の先生です。(メインの指導教員、メンターに次ぐ3人目という意味)
- Proposal の examiner (OIST 内部の先生 / 外部の先生): 2年次にproposalを審査していただく先生です。
- メインの指導教員 (Thesis Supervisor): ローテーション終了後から4年間所属する研究室のPIの先生を指します。
1年生
ローテーションと授業をこなします。
- 入学直後に、メンターの先生を誰にするかと、在学中に取る授業を決めます。メンターの先生を誰にするかについては、学生課から自分と分野の近い先生が紹介されるかと思います。後から変更することも可能です。
授業決めはメンターとしっかり相談してもいいし、特に相談しない人もいるようです。(制度上メールでメンターの了承を取る必要があります)
授業も後から変更できます。 - メンターとのmeeting(最低でも年に1回。3年次からはResearch Progress Reviewに切り替わる)
定期的にmeetingしてもらいます。メンターも時間がないので、要点をまとめて話をすると良いでしょう。meetingが終わったら、メンターの先生から学生課にレポートを提出していただきます。 - Rotation proposal と Rotation report
ローテーションプロポーザルではA4 1ページでこれからやることの提案書を書きます。各ローテーションが始まって1ヶ月が締め切りです。
ローテーションレポートはA4 3ページでローテーションでやったことをまとめたレポートを書きます。ローテーションが終わる月の最終日が締め切りです。分量は短いですが、練習だと思ってできるだけ良いものを作りましょう。 - おすすめの1年生の過ごし方
(2年生は結構忙しくなる人が多いので) 1年生のうちに予備的な研究成果を出したり、文献調査をたくさんしておくのが良いかと思います。また、運要素もありますが学振の業績欄に書くことを見越して、論文を書いて学術誌に投稿することにトライしてみましょう。
手法としては、
・メイン所属ラボで1年くらいで論文化できそうな小さいテーマに取り組む
・ローテーション先のラボで既に論文化しそうなプロジェクトを手伝って共著に入れてもらう
・OIST入学前の所属大学での卒論や修論を論文化する
等です。
いずれにしても早めに相談に行くにこしたことはないです
ちなみに、自分のメインラボをA研究室としたとき、ローテーションでB研究室に所属している際にも、空いた時間でA研究室に出入りして実験したりすることは可能です。(もちろん両ラボの先生に事前に許可を取っておく)
2年生
2年生での大きな目標はResearch proposal の提出 & Proposal exam の突破です。
単位が取りきれてない場合は授業も取りましょう。
ほとんどの学生は学振も書きます。
OIST学生はもともと大学から給料が支払われていますが、学振を取るとさらに給料が数万アップ + 特別研究員としての研究費を獲得することができます。
PCDという授業の一貫でグループプロジェクトというのもあります。
- PCD グループプロジェクト
入学年度の1月くらいに同学年の学生と3-5人(?) のグループを作ります。グループになったメンバーと協力し、数ヶ月かけて何か地域貢献系のプロジェクトをやります。僕は沖縄の高校生に向けた科学教室を開催しました。このカリキュラムはコロナウイルスの感染状況にかなり影響を受けるため、今後どのような形で行われるのかは不明です。 - Provisional thesis proposal title & scope (入学してから16ヶ月が締切)
4ヶ月後に締め切りが控えるプロポーザルの暫定タイトルとアブストラクトを書きます(実際に提出するプロポーザルは、ここから内容が変わっても問題ないです)。この書類には特に評価がつきませんが、プロポーザルの下準備になるのでしっかり書きましょう。 - 論文指導委員 (Thesis Committee) の決定
論文指導委員は、自分のPI+2人の先生からなる計3人の指導チームのことで、海外のPhD課程ではよくある制度のようです。今後ともこの3名の先生方にはお世話になることになります。
具体的には、
・メインの指導教員
・メンター
・Co-supervisor または Third Comittee Member
の3名の先生を指名します。この3名は、プロポーザルの審査員を指名するまでに決定し、承認して頂く必要があります。
Co-supervisor または 3rd committee memberについて。
あなたの研究についてアドバイスをくれる3人目の先生です。
異なる2つの研究室の要素を取り入れた研究を行いたい場合、メインの指導教員に加えて、さらにもう1人の先生にCo-supervisor (指導教員その2) になってもらうことができます。その場合は3rd committee memberとして誰かについてもらうことはなくなります。メインの指導教員は定年前でなければいけませんが、Co-supervisor や 3rd commitee memberに指名する先生には年齢制限がありません。 -
Thesis proposal (入学してから20ヶ月が締切)
在学中に行う研究について、研究計画書を提出します。プロジェクトの背景の包括的なレビュー、研究のゴール、3~5つの到達目標、それぞれの目標を達成するためのアプローチ、既に得ている予備的な結果、卒業までのタイムライン等で構成されています。分量に特に規定はないですが、20 ~ 数十ページくらいの書類になる人が多いと思います。
締切の前に(理想は1ヶ月ほど前)、論文指導委員の先生方にproposalの完成前原稿を提出し、チェックを受けてサインをもらいます。プロポーザルの提出には論文指導委員の先生方全員からサインをもらう必要があります。 -
プロポーザルの提出前後で、プロポーザルの審査をしていただく審査員の先生を決めます。
OIST内部の先生から1人、外部の先生から1人指名します。
外部の先生がメインの審査員になります。世界中の先生の中から、審査をしていただきたい先生に依頼をします。どの大学の先生でも基本的に大丈夫なはずですが、最低一人はPhD学生を卒業させている必要があります。この審査会をきっかけにコネクションが生まれたりします。
(指導教員から、指導教員の知り合いの先生を外部審査員に指名するように勧められるケースもあります)
ボランティアで審査をしていただく先生を探すことになるので、なかなか先生が見つからず、審査会の日程が後ろ倒しになる場合もあるようです。 -
Thesis proposal Examination (プロポーザル審査会) (入学してから24ヶ月が締切)
Thesis proposalに書いた内容に基づき、内部審査員、外部審査員、チェアの3名に向けてチョークトークを行います。(実際にはホワイトボードです。)
審査会に持ち込んで良いのは事前に提出したThesis proposal (と持ち込みたければ使い慣れたホワイトボードマーカー) のみで、パソコン等の持ち込みはできません。
審査会の日取りは、審査員の先生方のスケジュールに依存します。 -
実際に行ったプロポーザル審査会対策
僕の場合、審査会の日程が決まった時点で、実験は片手間でできるクローニングや細胞のお世話に切り替えました。
朝決めた時間に来て論文 (特に審査員の先生が過去に出版した論文) を読み、お昼過ぎの少し眠くなる時間帯に実験をしながら目を覚まし、夕食前後はセミナールームでチョークトークの練習をする、という生活をおくりました。ラボのみなさんを集めて練習会に付き合っていただいたりもしました。友人や3rd committeeの先生のラボの方にも発表を見てもらいました。(見ていただいた皆様、本当にありがとうございました)
zoomで自分の発表を録画して練習する方もいるようです。Examの準備に際して参考になったwebサイト
www.chem-station.comほとんどの場合、すぐに落ちて退学になることはなく、Pass (一発合格), minar revision (軽い修正), major revision (大幅な修正) のいずれかの結果が来るはずなので、それぞれの案内に従います。
-
学振を書く
OISTのカリキュラムに含まれてはいませんが、OIST学生も学振に申請することが強く推奨されています。内部で先輩採択者を交えた説明会があったり、学生課で採択された先輩の申請書を閲覧できたりします。
ちなみに、既に色んな人が書いていることですが、学振を書くにあたって個人的に大事だと思ったことをまとめた記事はこちらです。
入学時期によってプロポーザルを書くタイミングと学振を書くタイミングが被ったりします。
学振の学内締め切り:5月末
プロポーザルの締め切り
5月入学の人 → 書類:12月末、審査会:4月末
9月入学の人 → 書類:4月末、審査会:8月末
1月入学の人 → 書類:8月末、審査会:12月末
3年生から5年生
バリバリ研究を進めて論文を書きます。
外部の研究費にもじゃんじゃん申請します。
授業も取りたいのがあれば取ります。
(特にキャリア形成系のセミナーの案内はよく来ます)
1年に一度、論文指導委員 (Thesis Committee) の先生方とmeeting (Research Progress Review) を行います。
最後に博論を書いて審査会でプレゼンを行います。
- 年に一度の進捗報告会 (Research Progress Review)
1年間の進捗状況と今後の計画を発表して、論文指導委員の先生方(自分のPI+2人の先生からなる計3人の指導チーム)からアドバイスをもらう会です。学会発表の練習だと思って頑張りましょう。
-
論文指導委員の先生方とのmeetingについては次の記事に良くまとまっていますので一部引用させていただきます。(引用文中のTACというのがResearch Progress Reviewに対応しています。)
(引用始め)
TACは、研究者としての能力を審査される、というよりは自分のプロジェクトについて外部の専門家のディスカッションし、意見を貰う機会と認識しています。
TACでは、発表や質疑応答に対する評価やフィードバックだけでなく、プロジェクトの方向性、取捨選択、ストーリ構成、研究の意義、研究結果を論文にまとめるタイミングなど様々なアドバイスを複数の専門家の視点からもらえます。例えば、私の場合は、イギリスの大学や別の研究科の教授を含めた4人がTACメンバーで、1時間半〜2時間くらいの時間をかけて、次にどんな実験やったら面白そうか、とか、この研究室とコラボレーションしたらどうか、とか、この実験をするにはこの点に注意しないといけない、といった意見をいただきました。
TACメンバーの研究が、基礎か応用か、動物をモデルとして用いるか植物か、ドライな研究かウェットか、どこの国で研究をしているかによって、プロジェクトの方向性や売り込み方、ストーリ構成、考え方が少しづつ違っていて、幅広い意見をもらえるため、大変勉強になります。
また、自分の研究が一般的にどう評価されるかを知ることができ、何よりも、経験も知識も豊富な専門家たちに時間をいただいて、自分のプロジェクトについてディスカッションできるのはとても貴重な機会です。また、PIの指導方法等に問題がないかどうかもこのTACで相談することができ、問題が発生した場合は、TACメンバーが問題解決に働いてくれます。
(引用終わり) - 以下はOIST学生が実際に行う流れです。
meetingの締め切り日の1ヶ月以上前に、学生側から各先生に日程調整のメールを出します。日程が決定したら各先生に確認の連絡をし、学生課のweb formからも申請を行います。
要旨、イントロ、これまでに得た結果、今後のタイムラインを含めた数ページのレポートを書きます。これはmeetingが終わったら締め切りまでに学生課に提出することになります。
ほとんどの場合、発表にはパワポを使うので、それの準備もします。
meetingの10日以上前には、レポートとスライドをメインの指導教員に添削してもらうと良いと思います。
meetingの前日くらいになったら、リマインドも兼ねて各先生にレポートを送っておきます。
当日は発表をし、フィードバックをいただきます。
終了後に先生方には評価書を書いていただくので、評価書はここから提出して下さいね~ (提出先のlinkを添えて) のメールを送ります。
最後に、各先生に送ったレポートを学生課のweb formにも提出して終了です。
-
- 5年目の最後までに博論を書き、博論審査会をして合格したら卒業です。
以上5年間をまとめるとこんな感じです (OIST公式より引用)。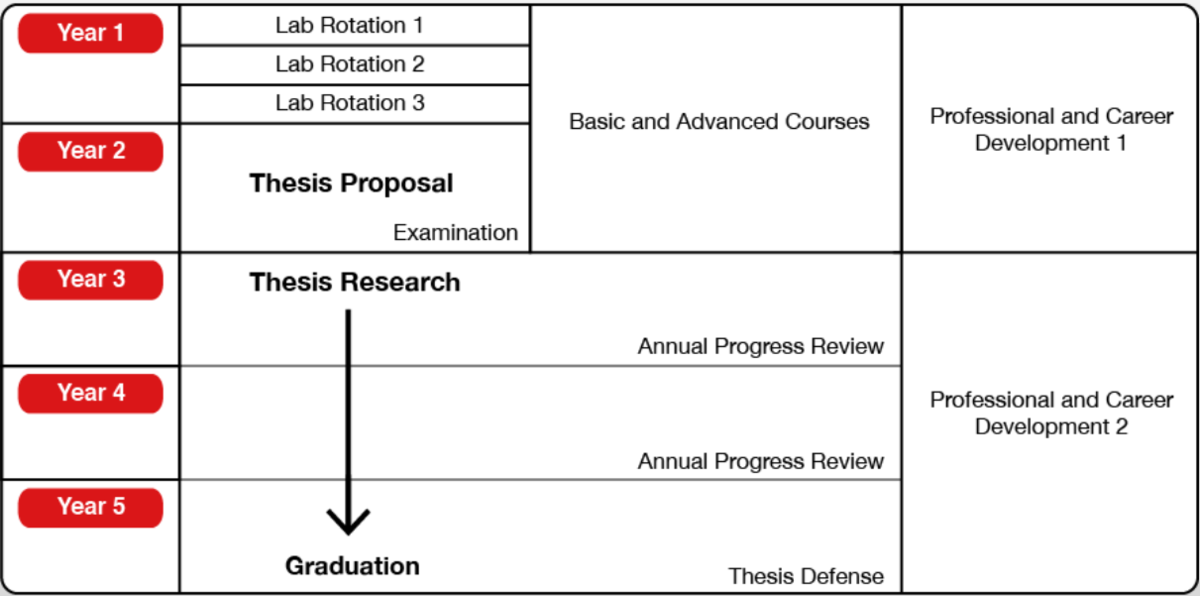
公式に載っていなさそうなQ&A
- OISTに入学したら最初の学振 (DC1) はいつ出す?
2年生の3月~4月に準備を始め、学内締め切りの5月末 - 6月頭に提出します。
5月, 9月, 1月のどの時期に入学していても同じです。
(これは、DC1の応募資格が、採用開始時の在学月数によって決まるからです。採用開始時に "一貫制の博士課程第3年次相当(在学月数24 ヶ月以上36 ヶ月未満)に在学する者" という規定です。募集要項(PD・DC2・DC1) | 特別研究員|日本学術振興会) - OISTで学生をしながら応募できる外部資金は?
次年度の4月1日時点でOISTの学生(かつ35歳未満)の場合、任意のタイミングで(入学して初年度でもいけるはず)、笹川研究助成に出すことが出来ます。締め切りは10月ごろ?
僕の専門に寄っててあれですが、他には以下のようなものがあります。これ以外にもあったら教えて下さい。
孫正義財団 (25歳以下)
戸部眞紀財団 (30歳以下)
ANRI (年齢制限無し)
日本ジェネティクス (生物系、35歳以下)
リカケンホールディングス (生物系、30歳以下)
ちなみにJST次世代や文科フェ(参考:こばしり on Twitter)はOISTの学生は対象外なので出せません。 - 締切に間に合わないとどうなる?
基本的にどんな締切も延長申請をすることで延長することが可能なはずです。ただし、就学年数は5年と決められており、それを過ぎると経済支援は打ち切られてしまうようです。 - 卒業要件は?
OISTとしては、論文の投稿まですれば卒業できるという話を聞いています (年度によって制度が改訂されるので要問合せ) 。また、卒業要件に独自の基準を設けている研究室もあるようです (最低2本は論文出してから卒業しようね~的な) 。
OISTは、学部卒で来た人も修士卒の人もほぼ同じ扱いを受けます (大きな違いは修士卒だと取らないといけない授業の単位数が少ないくらい) ので、学部卒からOISTに入学した人も、修士卒の人も、博士号にふさわしいと判断されれば3年くらいで卒業することは理論上可能だと思います。

